
Algal and Cyanobacterial Lectins and Their Antimicrobial Properties
by
 , , ,
, , ,
José Abel Fernández Romero
1,2,* ,
,
María Gabriela Paglini
3,4,
Christine Priano
1,
Adolfina Koroch
1,
Yoel Rodríguez
5,
James Sailer
2 and
Natalia Teleshova
2 1
Department of Science, City University of New York Borough of Manhattan Community College, New York, NY 10007, USA
2
Center for Biomedical Research, Population Council, New York, NY 10605, USA
3
Facultad de Ciencias Médicas, Instituto de Virología “Dr. JM Vanella”, Universidad Nacional de Córdoba, Córdoba X5000GYA, Argentina
4
Instituto de Investigación Médica Mercedes y Martín Ferreyra, INIMEC-CONICET-Universidad Nacional de Córdoba, Córdoba X5016GCT, Argentina
5
Department of Natural Sciences, City University of New York Hostos Community College, New York, NY 10451, USA
*
Author to whom correspondence should be addressed.
Mar. Drugs 2021, 19(12), 687; https://doi.org/10.3390/md19120687
Submission received: 2 November 2021
/
Revised: 19 November 2021
/
Accepted: 25 November 2021
/
Published: 1 December 2021
(This article belongs to the Special Issue Marine Lectins)
Abstract
:Lectins are proteins with a remarkably high affinity and specificity for carbohydrates. Many organisms naturally produce them, including animals, plants, fungi, protists, bacteria, archaea, and viruses. The present report focuses on lectins produced by marine or freshwater organisms, in particular algae and cyanobacteria. We explore their structure, function, classification, and antimicrobial properties. Furthermore, we look at the expression of lectins in heterologous systems and the current research on the preclinical and clinical evaluation of these fascinating molecules. The further development of these molecules might positively impact human health, particularly the prevention or treatment of diseases caused by pathogens such as human immunodeficiency virus, influenza, and severe acute respiratory coronaviruses, among others.
1. Introduction
1.1. Lectins, Their Structure, Function, and Carbohydrate-Binding Specificity
The Latin root for “lectin” means to choose or select, an appropriate meaning given that lectins are proteins that “choose”: to bind carbohydrates in glycolipids or glycoproteins and that the interaction of lectins with carbohydrates can be very selective and as specific as the antigen/antibody interactions. In addition to binding to oligosaccharides, they might also bind to monosaccharides, although with less affinity. Lectins are ubiquitous and can be produced by different organisms, including animals, plants, fungi, protists, and microorganisms such as bacteria, archaea, or viruses. There are different types of lectins shown in Table 1 that have been classified based on structural and functional similarities. The different types of lectins include C-type lectins (selectins, collectins, and endocytic lectins), S-type lectins (galectins), siglecs (sialic acid-binding Ig-like lectins), L-type lectins, P-type lectins, M-type lectins, Jacalin-related lectin (JRL), Cyanovirin-N homologs (CVNHs), Oscillatoria agardhii agglutinin homolog (OAAH), and Galanthus nivalis agglutinin-like (GNA-like) lectins, among others [1].
Each lectin molecule usually contains several binding sites for the simultaneous binding to multiple units of the carbohydrates they target. The three-dimensional structures of these proteins and their binding to carbohydrates can be very different among various lectins. For example, griffithsin (GRFT; Table 1) comprises 121 amino acids, where residue 31 does not appear to correspond to any standard amino acid [2]. GRFT adopts a β-prism I motif [3] observed in a variety of lectins and other proteins [4]. This motif consists of three repeats of a four-stranded antiparallel β-sheet forming a triangular prism [5]. Different from other related lectins, GRFT is defined by a domain-swapped homodimer where the first 2 β-strands of one monomer are linked to 10 β-strands of the other monomer, and vice versa [6]. Several X-ray crystal structures of GRFT in complex with monosaccharides and disaccharides have been solved. These include mannose (PDB IDs 2GUC, 2GUD, and 3LL2), N-acetylglucosamine (PDB ID 2GUE), 1→ 6α−mannobiose (PDB ID 2HYQ), and maltose (PDB ID 2HYR) [6,7,8]. For example, Figure 1 shows the complex between GRFT and six mannoses corresponding to six carbohydrate (mannose)-binding sites. Each GRFT mannose-binding site encloses Tyr and Asp residues in which both amino acids form three hydrogen bonds with each mannose. The three GRFT mannose-binding sites in each monomer are arranged to create an almost perfect equilateral triangle.
Similarly, cyanovirin-N (CV-N; Table 1) is an elongated lectin and consists of two similar domains, each with five β-strands and two helical turns. These two domains are formed after domain swapping and are connected by helical turns [5]. This domain swapping is present in both CV-N and GRFT; however, CV-N dimers swap half of the molecule, while in GRFT only 2 β-strands out of 12 are swapped. Additionally, CV-N has been naturally isolated in monomeric and dimeric forms; whereas other lectins, such as Microcystis viridis lectin (MVL; Table 1) and GRFT, are naturally produced only in their dimeric form. Furthermore, other lectins such as scytovirin (SVN; Table 1) seem to be produced in the monomeric form [5].
The role lectins play varies in the different organisms that produce them; they are involved in various biological processes. For example, in mammalians, lectins can be involved in cell-to-cell self-recognition, gamete fertilization, embryonic development, cell differentiation, apoptosis, immunomodulation, and inflammation, among other functions. In the case of marine or freshwater organisms, the main topic of this review, the functions that lectins play have been associated with cell-to-cell recognition, attachment, and bioflocculation, providing a competitive advantage over other organisms in their natural habitat [9].
1.2. Algal and Cyanobacterial Lectins: Their Classification and Characteristics
Marine species account for about half of total biodiversity and thus have received significant attention for their ability to produce natural molecules with potential biomedical properties [10,11]. One type of these molecules comprises lectins, proteins with a high affinity for carbohydrates, particularly those found on the surface of many pathogens. Significant sources of biomedical lectins include cyanobacteria (17%), green algae (22%), and red algae (61%) [12].
Cyanobacteria are a diverse group of oxygenic photosynthetic prokaryotes commonly found in a broad range of aquatic and terrestrial environments. They flourish at the surfaces of lakes and oceans and form mats in benthic environments. They can tolerate higher temperatures than eukaryotic cells, and can tolerate high salinity environments, desiccation, and water stress. Cyanobacteria are also known as blue-green algae because they produce phycocyanin pigment, which gives the cells a bluish color when present in high concentrations [13]. In addition to playing a pivotal role in changing the composition of the planet’s atmosphere, several species have been studied for their ability to produce biomolecules of importance in the biomedical field.
Marine algae are diverse organisms: Rhodophyta (red algae) and Chlorophyta (green algae) differ in their pigments, anatomy, and reproduction. They are one of the oldest types of marine organisms on earth and require salty/brackish water and sunlight. They are usually found attached to rocky surfaces. Like cyanobacteria, marine algae constitute an essential source of biomolecules, particularly lectins, that have promising applications in biomedicine [14]. Table 1 summarizes some of the most relevant lectins identified in these marine species.
2. Antiviral Activity of Marine Lectins
2.1. Mannose Arrays on Viral Spikes as a Target for Lectins
Glycans (oligo- and polysaccharides) shield protein surfaces of viruses, providing protection against the enzymatic activity of proteases. A diverse number of oligosaccharides can be displayed on the surface of mammalian cells, which are not a target for lectins that generally require repetitive arrays of specific sugars for recognition. However, repetitive arrays, such as high-mannose arrays, can be found on the surface of pathogens. For this reason, and because this mannose content is uncommon in mammalian cells, lectins are selective antimicrobial agents capable of binding the pathogen’s glycoproteins with high affinity and are not likely to be toxic to human cells [15,52,53,54].
To understand better the role of these glycans and how they might constitute an important target for antimicrobial agents and immune response, it is necessary to take a closer look at glycosylation in the HIV spike protein. The HIV-1 viral spike is one of the most heavily glycosylated proteins. It is challenging for the host humoral response to produce anti-glycan antibodies. The glycan network on an HIV-1 spike is often referred to as the glycan shield. The main function of these glycans is to protect conserved regions of the spike protein from neutralizing antibodies. Still, some neutralizing antibodies capable of binding to the glycans on an HIV-1 spike can be generated by the host humoral response. Interestingly, a cluster of high-mannose glycans that includes the N-linked glycan at position 332 can be targeted by different broadly neutralizing antibodies (bNAbs) [55]. The main glycan-dependent sites on the HIV-1 spike include not only the N332 glycan/V3 loop in the intrinsic mannose patch, but also the N160 glycan/V1/V2 loops and the glycans near gp120/gp41 interface [56]. Some of the broadly neutralizing antibodies that interact with these glycans include PGT128, PGT121, 10-1074, PGT135, PG9, PG16, PGT145, CAP256-VRC26.25, CH04, PGT151, 35O22, and 8ANC195 [57,58,59,60,61,62,63,64,65]. As a result, bNAbs have appropriately been the focus of significant research in preventing, treating, or even curing HIV.
Lectins can play a similar role when compared to bNAbs, and in many cases have a broader spectrum of neutralization. GRFT, CV-N, and SVN can bind to these glycans with high affinity and neutralize virions at a half-maximal effective concentration (EC50) as low as 30 pM. These lectins are not HIV subtype-specific because similar high-mannose arrays are present in all HIV subtypes. The 234 and 295 glycosylation sites are involved in GRFT, CV-N, and SVN neutralization of HIV-1 [52]. HIV can develop resistance to lectins by deglycosylation in site-specific residues (glycans at positions 234 and 295); however, multiple mutations are necessary [66]. Consequently, the deglycosylation required for resistance development might render a virus more susceptible to neutralization by the immune system [67].
2.1.1. Anti-HIV—Mode of Action and Efficacy Studies
HIV belongs to the viral family Retroviridae. It is an enveloped, positive-sense single-stranded RNA virus with the ability to reverse transcribe its genome and integrate the viral genome into the host genetic material. The viral reverse transcriptase controls the initial process of reverse transcription, and the lack of proofreading contributes to the high frequency of mutations that is clearly observed in the viral spike as a mechanism to evade humoral response. The initial process of viral replication starts with attachment to the CD4 receptor followed by interaction with a co-receptor, mainly CCR5 and CXCR4, and entry into a host cell. Many neutralizing molecules have been studied to block HIV entry into target cells. Because of the properties of lectins noted above, the potential for broad-spectrum efficacy, the high affinity, and the relatively low required concentration, lectins are increasingly being analyzed for their potential in this regard.
Recombinant GRFT has potent antiviral activity against HIV-1 strains and primary isolates with EC50 values in the low pM concentration. GRFT also interferes with cell-to-cell fusion and the transmission of HIV-1. GRFT blocks CD4-dependent glycoprotein (gp) 120 binding and inhibits gp120 binding of the monoclonal antibody (mAb) 2G12, which targets a carbohydrate-dependent motif, and the (mAb) 48d, that binds to CD4-induced epitope [2]. Mutations in the domain-swapped homodimer of GRFT, specifically in two or four amino acids at the dimerization interface, results in a monomeric form of GRFT. Cell-based assays indicate that this monomeric form of the lectin has greatly reduced activity against HIV-1, probably due to reduced crosslinking and aggregation of viral particles via multivalent interactions between GRFT and oligosaccharides present on HIV envelope glycoproteins [7]. Furthermore, GRFT, CV-N, and SVN are inhibitory for HIV-1 transfer from dendritic cells (DC-SIGN-mediated transfer of HIV-1) to CD4+ T cells, with GRFT being the most potent [68,69].
Different N-glycosylated sites have been implicated in mutations that affect the antiviral properties of lectins. Site-directed mutagenesis demonstrated that a single deglycosylation at N295 or N448 in HIV-1 isolates resulted in remarkable resistance to GRFT while still sensitive to CV-N, GNA, and some neutralizing antibodies. Unlike CV-N and GNA, the interaction between GRFT and gp120 appeared to be dependent on the specific trimer glycosylation, including N295 and N448 [67]. Deglycosylation of HIV-1 gp120 at position 295 or 448 decreases the sensitivity of transmitted/founder (T/F) gp120 to GRFT and at 339 to both CV-N and GNA. Mutation of all the three glycans renders a T/F gp120 resistant to GRFT [70]. HIV-1 subtype C develops resistance to GRFT, CV-N, and SVN after deletion of 1 to 5 mannose-rich glycans. Glycosylation sites at positions 230, 234, 241, and 289 located in the C2 region, and at positions 339, 392, and 448 in the C3-C4 region are involved in this resistance. The loss of glycosylation sites on gp120 and rearrangement of glycans in V4 are involved in HIV-1 subtype C resistance to GRFT, CV-N, and SVN [71].
MVN shares 33% identity with CV-N, and both lectins bind to similar carbohydrate structures. MVN and CV-N inhibit syncytium formation between HIV-1-infected T cells and uninfected CD4+ T cells and inhibit DC-SIGN-mediated HIV-1 binding and transmission to CD4+ T cells. HIV-1 exposure to dose-escalating concentrations of MVN selects a mutant virus with four deleted high-mannose-type glycans in gp120. The MVN-resistant virus is still sensitive to CV-N, HHA, GNA, and UDA but not to the 2G12 monoclonal antibody. Importantly, MVN is significantly less cytotoxic than CV-N [72].
Combinations of GRFT with tenofovir, maraviroc, and enfuvirtide showed synergistic activity against HIV-1 clade B and clade C isolates in primary peripheral blood mononuclear cells and in CD4+ MT-4 cells [20]. Similarly, paired carbohydrate-binding agent combinations show synergistic activity against HIV-1 and HIV-2, the 2G12 monoclonal antibody-resistant HIV-1, and MVN-resistant HIV-1. There is synergy in HHA and GNA combination against 2G12 monoclonal antibody-resistant HIV-1 and MVN-resistant HIV-1 strains. These carbohydrate-binding agents might have distinct binding patterns on the gp120 envelope and therefore might not interfere with each other’s glycan-binding sites on gp120 [73].
Few in vivo studies have been performed to test the efficacy of lectins against HIV-1. CV-N is effective against vaginal and rectal SHIV89.6P transmission [14]. Repeated administration of L. jensenii strain bioengineered to produce the protein CV-N in macaques led to a 63% reduction in transmission of SHIVSF162P3 after repeated challenge and in the peak viral loads in colonized macaques with breakthrough infection [14]. In the case of GRFT, only one study has shown the efficacy of this lectin against vaginal challenge in a macaque model. GRFT combined with carrageenan (CG) in a freeze-dried fast-dissolving insert (FDI) formulation protects rhesus macaques from a high dose vaginal SHIV SF162P3 challenge four hours after FDI insertion. The same formulation protects mice vaginally against HSV-2 and HPV pseudovirus [18].
2.1.2. Anti-HSV—Mode of Action and Efficacy Studies
The Herpesviridae family includes important pathogens associated with human diseases. This is the case of HSV-1 and -2 enveloped DNA viruses. The entry of HSV into the host cell involves several viral glycoproteins displayed on the virion’s surface. The HSV glycoprotein C (gC) and glycoprotein B (gB) bind to heparan sulfate on the cell surface to start the process of viral attachment to host cells. Next, glycoprotein D (gD) binds to entry receptors leading to subsequent steps with the participation of other viral glycoproteins such as glycoproteins H (gH) and L (gL). Ultimately, the virus fuses with the host cell membrane and enters the cell to replicate and produce new viral progeny [74].
The participation of viral glycoproteins, highly glycosylated, is fundamental for the successful attachment and entry to target cells. These glycoproteins have been described as potential targets for lectins, such as GRFT, that can interfere with viral entry. Levendosky and collaborators showed that GRFT might inhibit entry but not attachment to target cells [24]. Glycoprotein gD seems to be pivotal in explaining the mode of action of GRFT as this glycoprotein participates in both viral entry and cell-to-cell spread but not in viral adsorption. Moreover, gD N-glycosylation sites, potential targets for GRFT, are close to the gD functional domains for binding the gD receptor [75]. Additionally, Nixon and collaborators have proposed that GRFT might inhibit HSV-2 replication by interfering with cell-to-cell spread and by primarily targeting gB [29]. In this study, Nixon and collaborators showed that a 0.1% GRFT gel, but not placebo gel, significantly reduced HSV-2 disease scores in a murine model and resulted in greater survival even in the presence of a human seminal plasma [29]. GRFT combined with CG in a gel reduced HSV-2 vaginal infection in mice when given one hour before challenge with a high HSV-2 dose (106 plaque-forming units per mouse) [24]. Similarly, a GRFT and CG combination in a fast-dissolving insert (FDI) protected mice from HSV-2 infection four hours after FDI vaginal insertion [18].
2.1.3. Anti-HCV—Mode of Action and Efficacy Studies
Hepatitis C virus (HCV) is a member of the Flaviviridae family that causes acute and chronic liver inflammation, sometimes leading to serious liver damage and infecting humans and chimpanzees through direct blood contact. HCV is a positive-sense, single-stranded enveloped RNA virus of approximately 9600 nucleotides in length, depending on the genotype. There are seven main HCV genotypes (numbered 1–7) that vary by over 30% in nucleotide sequence [76,77,78]. The genome of HCV encodes for a single polyprotein precursor of about 3000 amino acids that is cleaved by both viral and cellular proteases to generate 10 mature viral proteins. These include four structural proteins: C (Core), E1 and E2 (envelope proteins 1 and 2) and p7 (which functions as a proton channel); and six non-structural proteins (NS2, NS3, NS4A, NS4B, NS5A, and NS5B) that have enzymatic and other activities necessary for virus replication [79,80,81,82]. E1 and E2 are the two envelope glycoproteins present on the surface of viral particles and essential for virus attachment to its receptor/co-receptors and for subsequent internalization into the target cells via a pH-dependent and clathrin-mediated endocytosis [83,84,85]. Each protein comprises a large N-terminal ectodomain responsible for virus attachment to its receptors, and a C-terminal transmembrane domain that anchors each glycoprotein in the lipid bilayer. The N-terminal ectodomains are strongly glycosylated, and the structure is stabilized by disulfide bridges [82,86]. Interestingly, one-third of the total molecular mass of the heterodimers E1-E2 corresponds to N-glycans that are highly conserved from most genotypes. More-over, these sites harbor high-mannose-type glycans, even after egress of viral particles from the cells [87]. These features, and the conserved nature of the entry steps of HCV, offer an attractive target for using lectins to develop strategies to prevent infection, especially in the context of liver transplants and for therapeutic intervention.
The progress in the development of retroviral particles pseudotyped with HCV envelope glycoproteins (HCVpp) and the amplification of HCV in cell culture (HCVcc) allow the identification and characterization of potential molecules that inhibit HCV entry to the host cell. In addition, it has been reported that the cyanobacterial lectins, CV-N, and MVL inhibit HCV entry by blocking the interaction between E2 and the receptor CD81 present in the membrane of the host cell. CV-N binds to high-mannose glycans (Man-8 and Man-9) in the E2 protein that forms part of the viral particle, exerting its inhibitory action at nanomolar concentrations in a dose-dependent manner. This was tested with different HCV genotypes in vitro using Huh-7 human hepatoma cells [11,88]. CV-N was found to be a more potent inhibitor than MVL, with EC50 concentrations ranging from 0.6 to 1.9 nM; whereas EC50 values of MVL ranged from 14.1 to 34.3 nM, more than 10-fold higher [89]. Another high-mannose-specific lectin, SVN, has also been shown to prevent HCV viral entry. In an infectivity assay using HCVpp bearing HCV E1 and E2 from various genotypes, SVN displayed inhibitory action with an EC50 of about 17 nM [32].
GRFT has been explored as a promising therapeutic antiviral strategy. GRFT exhibits a potent and broad-spectrum antiviral activity, with EC50 in the picomolar range. More-over, this lectin displays low in vitro and in vivo host toxicity and favorable subclinical outcomes range [90,91]. GRFT efficiently prevents HCV infection in cell culture (EC50 of 13.9 nM and EC90 of 49.5 nM), regardless of genotype, and it has been demonstrated that it also interferes with the direct cell-to-cell transmission of the virus (EC50 of 770 nM) [25]. The antiviral activity of GRFT lies in the ability to interact with the glycans present on the viral envelope proteins, E1-E2 heterodimer, thereby preventing the attachment of the virus to its receptor CD81 [25]. GRFT was successfully validated under in vivo conditions in chimeric mice that harbor primary human hepatocytes in their livers. Daily subcutaneous injections with 5 mg/kg GRFT could mitigate HCV infections. It is important to point out that GRFT can be safe and active when given on the same day as an HCV challenge [32]. The toxicity of GRTF in healthy mice was also evaluated, showing that treatments up to 50 mg/kg are well tolerated, with only mild toxicities [25,32].
The antiviral activity of all these lectins from red and blue-green algae against all genotypes of HCV can be explained by their targeting of high-mannose oligosaccharides on either E1 or E2 envelope proteins, which are common and conserved in all HCV genotypes.
2.1.4. Anti-Influenza Virus—Mode of Action and Efficacy Studies
Influenza viruses are enveloped, negative-sense single-stranded, segmented RNA viruses of the Orthomyxoviridae family that are classified into A, B, C, and D types based on antigen differences in their matrix and nucleoprotein. Human influenza A and B viruses are highly contagious and are primarily responsible for the seasonally acute respiratory disease that outbreaks and spreads worldwide, giving rise to increased hospital admission and mortality. However, attention is focused mainly on influenza type A because it is by far the most virulent and causes severe respiratory disease or death [92,93].
Each virion contains three main components: a viral envelope of a lipid bilayer is decorated with three transmembrane proteins, namely hemagglutinin (HA), neuraminidase (NA), and an ion channel (M2); an intermediate layer of matrix protein (M1); and the third innermost component, the helical viral ribonucleocapsid (vRNP) core formed by the nucleoprotein (NP) and the eight segments of single-stranded negative-sense RNA (vRNA) [94]. The surface of the influenza A virus is covered with a large number of glycoproteins HA and NA, which are the basis of further subdivisions of the virus. To date, 18 subtypes of HA and 11 subtypes of NA have been identified by The United States Centers for Disease Control and Prevention (CDC); however, only H1-3 and N1-2 strains have achieved human-to-human transmission and are currently circulating as seasonal influenza strains (https://www.cdc.gov/flu/about/viruses/types.htm, accessed on 26 November 2021).
HA represents 80% of the surface viral glycoproteins in a ratio HA/NA of about 300/40, influenced by the genetic background and subtype of the virus [95]. These two glycoproteins, HA and NA, recognize the same molecule located at the host cell membrane’s surface, the N-acetylneuraminic (sialic) acid (SA) receptor [96]. HA plays two crucial roles during infection. First, it binds to the terminal SA on oligosaccharide chains of glycoproteins or glycolipids present at the host cell membrane, the first stage of viral infection. Further, it allows the fusion of viral and endosomal membranes, thereby delivering the viral nucleocapsid into the cytoplasm to initiate viral replication [97]. HA is a trimer of indistinguishable subunits, each one containing a receptor-binding site highly conserved in all subtypes of influenza [96]. NA is a tetramer of four identical polypeptide chains. This protein is a glycoside hydrolase mediating hydrolysis of the link between the SA bound to HA, relevant in promoting the release of new virus and preventing their aggregation on the host cell surface [98]. Thus, HA and NA have opposing roles, and the balance between their activities can be crucial in securing the infectivity of a particular viral strain. The functional and crucial role of HA in viral attachment and membrane fusion makes it an interesting target for antiviral drugs. In this regard, HA carries potential sites for N-linked glycosylation, is highly conserved between the different subtypes, and contributes to the stability of the molecule [99]. Hence, agents that interact with the glycans of HA to inhibit entry of the virus into susceptible target cells are excellent candidates.
Among potential agents, therefore, lectins that specifically recognize carbohydrate (glycan) structures are especially attractive for the development of prophylactic or therapeutic treatments of influenza virus infection. CV-N is highly active against a wide spectrum of influenza A and B virus strains, including clinical isolates and an in vitro-derived neuraminidase inhibitor-resistant strain. It exhibits antiviral activity by binding to high-mannose (oligomannose-8 and oligomannose-9) residues on HA1 molecules, resulting in virus neutralization with EC50 values from 0.004 to 0.5 g/mL, as tested in MDCK cells. However, there are two strains of the influenza virus, A/PR/8/34 (H1N1) and NWS/33 (H1N1), that are insensitive to CV-N even at concentrations of 10 g/mL. Loss of glycosylation sites of HA due to mutations leads to decreases in CV-N binding and antiviral efficacy [39,40,42]. The activity of CV-N was also assayed in animal models. In mice and ferrets, CV-N efficacy was dose-responsive from 0.0625 to 1 mg/kg/day when administered intranasally twice daily before viral infection. A significant reduction in viral titers in the lungs of mice and nasal washes in ferrets were observed [41]. However, CV-N has drawbacks in pharmaceutical applications, such as a short plasma half-life, proteolysis, and immunogenicity. PEGylation of CV-N as well as the linker-CV-N (LCVN) improved its pharmacokinetic and pharmacodynamic properties and maintained high affinity toward glycans containing mannose linkages [100]. These compounds, PEG20k-LCVN and LCVN, exhibited potent and selective anti-influenza activity in nanomolar concentrations in infected MDCK cells. In chicken embryos, they completely inhibited H3N2 virus propagation (strain A/HK/8/68) in micromolar concentrations [40,100].
The lectin BCA showed potent anti-influenza activity against most influenza virus strains in MDCK cells, including a clinical isolate of pandemic H1N1-2009 virus; however, the sensitivity was much lower (EC50 of 800 nM) compared with other strains [12,49].
Another two high-mannose-binding lectins, KAA-2 and HRL40, showed strong antiviral activity against influenza virus in MDCK and NCI-H292 cells, respectively. The activity was regardless of the virus strain and subtype, with EC50 in the low nanomolar levels. KAA-2 bound exclusively to high-mannose-type N-glycans, present at the globular head domain of HA, inhibiting the entry of the virus into the cell [12,47]. Similarly, HRL40 showed a potent anti-influenza activity through high-affinity binding to the viral hemagglutinin spike [50].
2.1.5. Anti-EBOV—Mode of Action and Efficacy Studies
EBOV is the most prominent member of the Filoviridae family that includes enveloped viruses with a single-stranded, negative-sense RNA genome. Ebola virus disease (EBOVD) is a rare and deadly disease caused by infection with the EBOV. EBOV causes severe hemorrhagic fever in humans and nonhuman primates, with human case fatality rates of up to 90%. The virus first spreads to people through direct contact with the blood, body fluids, and tissues of animals. EBOV then spreads to other people through direct contact with the body fluids of a sick person or someone who has died from EBOVD [101,102].
The EBOV genome is a linear, non-segmented, single-stranded RNA of approximately 19 kb. The viral genome encodes seven proteins, including a nucleoprotein (NP), a glycoprotein (GP), an RNA-dependent RNA polymerase (L), and four structural proteins termed VP24, VP30, VP35, and VP40. Structural proteins VP40 and VP24 conform to the viral matrix linking the nucleocapsid to the viral envelope. GP is an integral membrane protein of the viral envelope that forms spike-like protrusions on the surface of the virion and is involved in the internalization process to the host cell [103,104]. GP is cleaved by furin into disulfide-linked GP1 and GP2 subunits [105,106]. GP1 is responsible for cellular attachment, while GP2 mediates viral and host membranes fusion [107]. The GP1 subunit contains the putative receptor-binding region and a heavily glycosylated region named mucin-like domain. GP1/2 molecule is a chalice-shaped trimer with a thick coating of oligosaccharides on a novel glycan cap and projecting mucin-like domain. The glycan cap and the mucin-like domain concentrate most of the N-linked glycosylation sites (including high-mannose oligosaccharides) and O-linked carbohydrates [107,108]. Thus, EBOV-GP is the main target for entry inhibitors, where lectins are excellent candidates. CV-N and SVN bind with high affinity to mannose-rich oligosaccharides on GP, blocking entry into target cells [51]. Antiviral activity of the lectins results from specific binding to the mucin region of GP, enriched in N-linked high-mannose oligosaccharides [109]. CV-N and SVN inhibit Zaire Ebola virus (EBOZV) infection, both in vitro and in vivo, through their ability to bind to oligomannoses-8/9 on the GP [36,51]. The use of recombinant systems to generate pseudotyped virus vectors carrying native GP or/and eGFP molecules has provided a powerful tool for studying the antiviral activity of several compounds. CV-N and SVN inhibit the replication of EBOZV-eGFP in Vero E6 cells with an EC50 of 154 nM and 41 nM [51]. Similar experiments using HeLa cells show that CV-N inhibits EBOZV GP-pseudotyped at lower concentrations (EC50 ~ 40–60 nM) [36]. On the other hand, when CV-N is used in vivo to treat EBOZV infected mice, repeated daily subcutaneous injections of approximately 5 mg/kg reduced the viral titers, and death was delayed but not prevented [37]. In vivo studies using SVN concluded that 90% of the mice receiving 30 mg/kg/day of SVN beginning on the day before virus inoculation survived the infection. Lower doses of SVN of 20 or 10 mg/kg/day showed 80% and 30% survival rates, respectively. Although SVN was highly protective in ZEBOV-infected mice, its short serum persistence required dosing every six hours [51], which is a disadvantage for any potential human therapy.
2.1.6. Anti-Coronaviruses—Mode of Action and Efficacy Studies
The Coronaviridae family includes notable zoonotic pathogens, including MERS, SARS-CoV-1, and SARS-CoV-2. These viruses are enveloped viruses with a positive-strand RNA genome and highly N-glycosylated spike proteins that mediate attachment and entry to the epithelial cells that line the respiratory tract in humans. Different lectins have shown the potential to target coronaviruses [110]. O’Keefe and collaborators first reported that GRFT binds to the SARS-CoV-1 spike glycoprotein and inhibits viral entry. They showed the activity of GRFT against a variety of SARS-CoV-1 strains, other human coronaviruses, and coronaviruses that infect other animal species. Mice treated with GRFT and exposed to a mouse-adapted SARS-CoV-1 showed significantly reduced morbidity and mortality and inhibited cytokines associated with deleterious aspects of the host immunological response to the infection [30].
The combination of GRFT with ι-CG or λ-CG results in synergistic antiviral activity using a pseudoviral model. While ι-CG or λ-CG alone inhibits the entry of SARS-CoV-1 and SARS-CoV-2 pseudoviruses with EC50 values between 3.2 and 7.5 µg/mL, combination with GRFT results in 10 times lower EC50 values. This synergistic activity was observed with different pseudoviral particles, including those containing recently identified mutations in the SARS-CoV-2 spike protein (D614G, K1417N/E484K/N501Y) [16]. GRFT has also shown a synergistic effect when combined with a pan-coronavirus fusion inhibitor (EK1) that targets the SARS-CoV-2 spike S2 subunit [111]. Using particles pseudotyped with the MERS-CoV spike protein, Millet and collaborators showed that GRFT interferes with MERS-CoV entry [112].
2.1.7. Anti-HPV—Mode of Action and Efficacy Studies
HPVs are small double-stranded DNA viruses in the Papovaviridae family with tropism for the skin and mucosal epithelia. Different from the other viruses discussed above, HPV is a naked virus, and the viral spikes are not glycosylated. Obviously, the virion per se cannot be targeted by lectins, but a unique mode of action might allow GRFT to interfere with HPV replication. Levendosky and collaborators proposed that GRFT might promote the internalization of the HPV secondary receptor in vitro. Additionally, the combination of GRFT and CG results in a potent anti-HPV activity in a murine model using HPV-16 pseudoviruses [18,24].
3. Antibacterial Activity of Marine and Freshwater Lectins
Though there are several examples in the literature of algal lectins as antivirals and several reports of lectins from many different sources that have antibacterial activity, there are relatively few studies in the literature that specifically address the potential usefulness of algal or cyanobacterial lectins as antibacterial agents [40,113,114]. This section will review the relevant information available. In all cases, standard microbiological methods were employed by researchers to test for inhibition of bacterial growth, including disk diffusion and culture density assays.
In one study, Liao et al. [115] showed that purified lectins isolated from two red algal species, Eucheuma serra (ESA) and Galaxaura marginate (GMA), strongly inhibited the growth of the pathogenic marine Gram-negative Vibrio vulnificus, although it showed no activity against two other Vibrio species, V. peagius and V. neresis. Selective inhibition was attributed to differences in bacterial surface carbohydrates. In addition, this study showed that whereas saline and ethanol extracts of several algal species exhibited antibacterial activity against both V. vulnicius and V. peaguius, this activity was inhibited by pre-treatment with lectin-binding sugars and glycoproteins, suggesting that lectins present in the algal extracts were the active inhibitory agents. Hung et al. [116] further demonstrated that lectins isolated from the red algae Eucheuma denitculatum (EDA) exhibit activity against another pathogenic marine vibrio, V. alginolyticus, but not against V. parahaemolyticus or V. harveyi. Binding assays suggested that selective activity was through binding of the EDA lectins to high-mannose N-glycans.
Holanda and colleagues reported differential activity of lectins from the marine red alga S. filiformis against human pathogenic bacteria [117]. In this study, isolated lectins inhibited the growth of Gram-negative bacteria Salmonella typhi, Serratia marcescens, Enterobacter aerogenes, Klebsiella pneumoniae, Pseudomonas aeruginosa, and Proteus sp. There was no inhibitory effect, however, on Gram-negative S. typhimurium or Escherichia coli, nor was there any inhibition of Gram-positive Staphylococcus aureus, Bacillus subtilis, or Bacillus cereus. For inhibited strains, bacterial cell density was significantly reduced in the presence of lectin at high concentrations (1 mg/mL), but cell density assays showed that bacteria exhibited all phases of cell growth. The investigators suggested that mannan present on cell walls of Gram-negative bacteria might bind to lectin and alter the flow of nutrients, thereby causing inhibition of growth [45,113].
Vasconcelos et al. tested the effect of isolated lectins from two species of red algae, Bryothamnion seaforthii (BSL) and Hypnea musciformis (HML), for their ability to inhibit growth in biofilms of Gram-positive S. epidermidis and S. aureus, and of Gram-negative Klebsiella oxytoca, P. aeruginosa, Candida albicans, and Candida tropicalis. Whereas HML and BSL both caused weak growth reductions in S. aureus, S. epidermidis, and P. aeruginosa, only HML reduced the growth of K. oxytoca [118]. For S. aureus, BSL decreased the biofilm mass at all concentrations used; however, HML caused only a small decrease at the highest concentration tested (250 µg/mL). In addition, these lectins caused only a small decrease in the number of viable S. aureus cells. The biofilm mass of K. oxytoca was also reduced in the presence of these lectins, but with no decrease in the number of viable cells [118].
Collectively, these investigators have suggested the potential usefulness of marine lectins as natural alternatives to conventional antibiotics in therapeutic interventions for infections by Gram-negative pathogens [117] and for the protection of marine species susceptible to marine vibrio infection [40,113,114]. With the global rise in antibiotic resistance over the past century, there has been a pressing need to find alternative sources of antibiotic agents for therapeutic use. As demonstrated in the studies reviewed here, the ability of algal lectins to inhibit the growth of various pathogenic bacteria makes them potential candidates for medicinal use and is worth further investigation.
4. Antiprotozoal Activity of Marine Lectins
Although metabolite extracts obtained from some marine algae have been shown to exhibit antiprotozoal activity, there are few reports in the literature of activity specifically attributed to algal or cyanobacterial lectins. Reports include the activity of algal extracts from Bostrychia tenella against Trypanosoma cruzi and Leishmania amazonensis [119]; from various species of brown algae against T. cruzi, Trypanosoma brucei rhodesiense, L. donovani, Sargassaceae sp. [120]; and from Anadyomene saldanhae, Caulerpa cupressoides, Canistrocarpus cervicornis, Dictyota sp., Ochtodes secundiramea, and Padina sp. against L. braziliensis. Chatterjee et al. further report that GRFT exhibits anti-protozoal activity against T. vaginalis and Tritichomonas foetus in a vaginal mouse model [17]. This is the only report of an algal lectin having such activity. Investigation into the possible activity of other marine lectins might yield novel insights into therapeutic uses against parasitic protozoan infections.
5. Expression of Marine Lectins in Heterologous Systems
Expressing lectins in heterologous systems can lead to a cost-effective production for pharmaceutical purposes. The heterologous systems provide higher yields than conventional purification and reduce the production cost and time [121]. Several models have been used for the heterologous production of lectins, such as bacteria, yeast, plants, mammalian, and insect cells. Figure 2 shows different strategies used for the expression and purification of GRFT and their intended use.
GRFT is the marine lectin in which expression in heterologous systems has been more widely explored. The expression of GRFT in tobacco plants (Nicotiana benthamiana) has been one of the choices. For this purpose, a synthetic cDNA (GenBank no. FJ594069) encoding the 121 amino acids of GRFT has been cloned into a tobacco mosaic virus (TMV) vector in which GRFT is expressed under the control of a duplicated coat protein subgenomic promoter. N. benthamiana seedlings are inoculated with infectious recombinant TMV, and the infected leaf biomass is processed 12 days after infection to extract GRFT [31]. The studies have shown that GRFT becomes the most abundant protein in the plant material and can be purified through filtration and chromatography. Through this process, GRFT accumulates at more than 1 g of GRFT per kilogram of N. benthamiana leaf material, providing a final product at concentrations above 20 mg/mL, and allowing production of more than 60 g of pure GRFT in a single 5000-square-foot enclosed greenhouse. The manufacturing cost of GRFT using this plant-based system has been estimated to be USD $0.32/dose. This assumes a commercial launch volume of 20 kg GRFT/year for 6.7 million doses of GRFT at 3 mg/dose, a recovery efficiency of 70%, and purity of >99%. This manufacturing process was also found to have a favorable environmental output with minimal risks to health and safety [122]. Additionally, gene-silencing suppressors for high-level production of GRFT in N. benthamiana have resulted in a higher accumulation of GRFT with a yield of 400 μg g−1 fresh weight or 287 μg g−1 after purification, representing a recovery of 71.75% [123].
Similarly, Nicotiana tabacum has been transformed with a vector containing the gene encoding CV-N. Through this process, the plant-derived CV-N can be recovered at 130 ng per mg of fresh leaf tissue. CV-N is expressed in the desired monomeric form using this plant-based system. Hydroponic culturing of transgenic plants results in CV-N rhizosecretion at 0.64 mg/mL hydroponic media after 24 days [124]. The transplastomic plants allow a highly efficient and cost-effective production platform for lectins, and dried tobacco can serve as a source material for the purification of lectins [125].
Oryza sativa (rice plant) has been used to express GRFT. For this purpose, GRFT has been expressed in the endosperm of transgenic rice plants. The yield of GRFT in this system can reach 223 g/g dry seed weight, and through a one-step purification protocol, can achieve a recovery of 74% and a purity of 80% [126].
Other host organisms for the economical and efficient production of lectins include bacteria. Hexa-histidine-tagged GRFT (His-GRFT) has been successfully produced in E. coli. Production in a fermenter with an auto-inducing medium allows the total amount of His-GRFT per liter to be increased by about 45-fold [127]. Similarly, recombinant expression in engineered E. coli results in GRFT concentrations of 2.5 g/L. This could translate into production volumes of >20 tons per year at the cost of goods sold below USD $3500/kg [128].
Finally, probiotic lactobacilli have been studied for the expression of lectins. Lactobacillus rhamnosus GG and GR-1 have been engineered to express GRFT [129], while Lactobacillus jensenii and Lactobacillus plantarum have been used to express CV-N [130,131,132] and SVN [133], respectively. The idea behind this strategy is that lectin-producing lactobacilli could colonize the mucosal epithelium and produce the lectin in vivo to protect the host if exposed to HIV or other pathogens.
6. Preclinical and Clinical Safety Studies of Marine and Freshwater Lectins
In addition to the potent antimicrobial activity, the safety of potential lectin-based products is of paramount importance. Comprehensive preclinical and clinical safety evaluations must be performed as part of product development [134]. This section reviews the preclinical and clinical studies that have been performed for these lectins.
CV-N: CV-N has been of particular interest for the development of a topical anti-HIV microbicide. Prolonged production of recombinant CV-N by L. jensenii vaginally in nonhuman primates did not induce any observable adverse effects or inflammatory biomarkers [135]. However, there are some safety concerns over the use of CV-N. CV-N affected PBMCs morphology, induced mitogenic activity in PBMCs, and increased expression of cellular activation markers and several cytokines following prolonged exposure [136,137]. CV-N can bind to cellular proteins and might induce potential toxic effects [89]. Furthermore, some paradoxical effects of CV-N enhancing R5 HIV infection at low concentrations were reported [138]. CV-N has been modified by site-specific conjugation with polyethylene glycol in a reaction called PEGylation to improve the drug-like properties of this lectin [139]. When administered intravenously, the PEGylated CV-N was significantly less immunogenic than CV-N [139]. The safety concerns observed in vitro have reduced the enthusiasm for this lectin.
MVN and MVL: MVN isolated from Microcystis aeruginosa shares partial homology with CV-N, has a potent but narrow anti-HIV profile, and demonstrates a better safety profile than CV-N [72]. However, several cytokines were significantly increased in PBMCs after exposure to this lectin [72]. The incubation of PBMCs with MVN leads to weak induction of expression of activation markers but did not activate or enhance viral replication in pretreated cells [72,140].
Like CV-N, MVL binds to the target cell surface and the viral envelope [89]. Cytotoxic effects triggered by the lectin might occur because of MVL interaction with cellular proteins. Indeed, MVL inhibited cell viability in several cell lines, including Hep-G2 (human hepatocellular liver carcinoma), HT-29 (human colon cancer), SGC-7901 (stomach cancer), and SK-OV-3 (human ovarian cancer) (IC50 40–53 µg/mL)) [141].
OAA: OAA is a stable protein [142]. However, the development of OAA-based products might be problematic because it exerts cytotoxic effects such as CV-N, MVN, and MVL [44].
GRFT: GRFT has an excellent safety profile. In contrast to CV-N, GRFT, with its broad and potent antiviral activity, does not have stimulatory properties [143]. The lectin inhibits HIV infection in human cervical explant tissues with no proinflammatory cytokine production. GRFT has an excellent safety profile when tested in a rabbit vaginal irritancy model [31] or when administered in single or chronic subcutaneous doses in mice and guinea pigs [144]. GRFT is safe and minimally absorbed after repeated vaginal application. Repeated dosing of GRFT and GRFT/CG gel in small animal models revealed no adverse findings at any dose levels tested and showed that a GRFT/CG gel is non-irritating. Seven days of daily vaginal application of 0.1% GRFT/CG gel did not enhance the susceptibility of mice to HSV-2 infection. Fourteen days of daily intravenous administration of GRFT up to 8.3 mg/kg/day in rats resulted in no detectable anti-drug-antibodies (ADA) and a no adverse effect level (NOAEL) of 8.3 mg/kg/day despite high systemic levels of GRFT. Fourteen days of daily vaginal GRFT/CG gel dosing (up to 0.3% GRFT) in rats resulted in a NOAEL of 0.3% GRFT and little or no vaginal irritation. This regimen also resulted in little or no systemic detection of GRFT. A related study in rabbits also found a NOAEL of 0.3% GRFT and little or no vaginal irritation [18].
GRFT’s preclinical safety, lack of systemic absorption after vaginal administration in animal studies, and lack of cross-resistance with existing antiretroviral drugs prompted its development for topical HIV pre-exposure prophylaxis (PrEP). The Population Council investigated the safety, pharmacokinetics, pharmacodynamics, and immunogenicity of a vaginal gel (PC-6500: 0.1% GRFT in a CG gel) in healthy women after vaginal administration. In this first-in-human trial of GRFT, no significant adverse events were recorded in clinical or laboratory results or histopathological evaluations in cervicovaginal mucosa. Additionally, no anti-drug (GRFT) antibodies were detected in serum, and no cervicovaginal proinflammatory responses or changes in the ectocervical transcriptome were evident. Decreased levels of proinflammatory chemokines in CVLs were observed, while GRFT was not detected in plasma after vaginal application. GRFT and GRFT/CG in CVL samples inhibited HIV and HPV, respectively, ex vivo in a dose-dependent manner. This study suggested that GRFT formulated in combination with CG is a safe and promising multipurpose prevention technology product that warrants further investigation (Teleshova et al. submitted). The Population Council is currently investigating fast-dissolving inserts and vaginal rings containing Q-GRFT alone or in combination with CG. Q-GRFT is a version of GRFT in which a methionine has been substituted by another amino acid (M78Q) to reduce the potential oxidation of this lectin. An additional phase 1 clinical trial (PREVENT), led by the University of Louisville and the University of Pittsburgh, was planned to look at a GRFT-based rectal microbicide safety. This trial was terminated, and the study enrollment was prematurely halted due to the COVID-19 pandemic. The trial was designed to study if a single dose of an enema containing Q-GRFT was safe, well tolerated, and acceptable in healthy adults practicing receptive anal intercourse [145].
Limited preclinical safety data are available for several promising lectins, including SVN, BCA, KAA-2, and HRL40.
7. Conclusions
The sui generis mode of action of mannose-binding lectins such as GRFT, SVN, CV-N, OAA, and MVL against important pathogens has prompted the development of these molecules as potential therapeutics or prophylactic drugs to target single or multiple infectious diseases. Lectins are highly specific, might show broad-spectrum activity, are locally (topically) delivered, and are relatively potent. As such, they deserve additional investigation. That said, there are safety concerns with some lectins that induce mitogenic activity after prolonged exposures, and mass production of lectins is a relatively nascent field. GRFT might be an especially promising candidate. GRFT has activity that seems to be among the broadest of any yet-evaluated lectin, is an exception to the mitogenic activity, has been tested in phase 1 clinical trials with promising results regarding its safety in topical formulations, and appears to be producible in cost-effective doses.
Author Contributions
Conceptualization, J.A.F.R.; visualization, Y.R. and J.A.F.R.; writing—original draft preparation J.A.F.R., M.G.P., C.P., A.K., Y.R., J.S. and N.T.; writing—review and editing J.A.F.R., M.G.P., C.P., A.K., Y.R., J.S. and N.T.; funding acquisition J.S. and J.A.F.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by The Population Council.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ghazarian, H.; Idoni, B.; Oppenheimer, S.B. A glycobiology review: Carbohydrates, lectins and implications in cancer therapeutics. Acta Histochem. 2011, 113, 236–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, T.; O’Keefe, B.R.; Sowder, R.C., 2nd; Bringans, S.; Gardella, R.; Berg, S.; Cochran, P.; Turpin, J.A.; Buckheit, R.W., Jr.; McMahon, J.B.; et al. Isolation and characterization of griffithsin, a novel HIV-inactivating protein, from the red alga Griffithsia sp. J. Biol. Chem. 2005, 280, 9345–9353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chothia, C.; Murzin, A.G. New folds for all-beta proteins. Structure 1993, 1, 217–222. [Google Scholar] [CrossRef]
- Shimizu, T.; Morikawa, K. The beta-prism: A new folding motif. Trends Biochem. Sci. 1996, 21, 3–6. [Google Scholar] [CrossRef]
- Ziolkowska, N.E.; Wlodawer, A. Structural studies of algal lectins with anti-HIV activity. Acta Biochim. Pol. 2006, 53, 617–626. [Google Scholar] [CrossRef] [Green Version]
- Ziolkowska, N.E.; O’Keefe, B.R.; Mori, T.; Zhu, C.; Giomarelli, B.; Vojdani, F.; Palmer, K.E.; McMahon, J.B.; Wlodawer, A. Domain-swapped structure of the potent antiviral protein griffithsin and its mode of carbohydrate binding. Structure 2006, 14, 1127–1135. [Google Scholar] [CrossRef] [Green Version]
- Moulaei, T.; Shenoy, S.R.; Giomarelli, B.; Thomas, C.; McMahon, J.B.; Dauter, Z.; O’Keefe, B.R.; Wlodawer, A. Monomerization of viral entry inhibitor griffithsin elucidates the relationship between multivalent binding to carbohydrates and anti-HIV activity. Structure 2010, 18, 1104–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziolkowska, N.E.; Shenoy, S.R.; O’Keefe, B.R.; McMahon, J.B.; Palmer, K.E.; Dwek, R.A.; Wormald, M.R.; Wlodawer, A. Crystallographic, thermodynamic, and molecular modeling studies of the mode of binding of oligosaccharides to the potent antiviral protein griffithsin. Proteins 2007, 67, 661–670. [Google Scholar] [CrossRef]
- Kotecha, H.; Poduval, P.B. Chapter 9—Microbial lectins: Roles and applications. In Advances in Biological Science Research: A Practical Approach; Academic Press: Cambridge, MA, USA, 2019; pp. 135–147. [Google Scholar]
- Aneiros, A.; Garateix, A. Bioactive peptides from marine sources: Pharmacological properties and isolation procedures. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2004, 803, 41–53. [Google Scholar] [CrossRef]
- Cheung, R.C.; Wong, J.H.; Pan, W.; Chan, Y.S.; Yin, C.; Dan, X.; Ng, T.B. Marine lectins and their medicinal applications. Appl. Microbiol. Biotechnol. 2015, 99, 3755–3773. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.S.; Thakur, S.R.; Bansal, P. Algal lectins as promising biomolecules for biomedical research. Crit. Rev. Microbiol. 2015, 41, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Cohen, Y.; Gurevitz, M. The Cyanobacteria—Ecology, physiology and molecular genetics. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006. [Google Scholar]
- Pagarete, A.; Ramos, A.S.; Puntervoll, P.; Allen, M.J.; Verdelho, V. Antiviral potential of algal metabolites—A comprehensive review. Mar. Drugs 2021, 19, 94. [Google Scholar] [CrossRef] [PubMed]
- Alexandre, K.B.; Gray, E.S.; Pantophlet, R.; Moore, P.L.; McMahon, J.B.; Chakauya, E.; O’Keefe, B.R.; Chikwamba, R.; Morris, L. Binding of the mannose-specific lectin, griffithsin, to HIV-1 gp120 exposes the CD4-binding site. J. Virol. 2011, 85, 9039–9050. [Google Scholar] [CrossRef] [Green Version]
- Alsaidi, S.; Cornejal, N.; Mahoney, O.; Melo, C.; Verma, N.; Bonnaire, T.; Chang, T.; O’Keefe, B.R.; Sailer, J.; Zydowsky, T.M.; et al. Griffithsin and carrageenan combination results in antiviral synergy against SARS-CoV-1 and 2 in a pseudoviral model. Mar. Drugs 2021, 19, 418. [Google Scholar] [CrossRef]
- Chatterjee, A.; Ratner, D.M.; Ryan, C.M.; Johnson, P.J.; O’Keefe, B.R.; Secor, W.E.; Anderson, D.J.; Robbins, P.W.; Samuelson, J. Anti-retroviral lectins have modest effects on adherence of trichomonas vaginalis to epithelial cells in vitro and on recovery of tritrichomonas foetus in a Mouse Vaginal Model. PLoS ONE 2015, 10, e0135340. [Google Scholar]
- Derby, N.; Lal, M.; Aravantinou, M.; Kizima, L.; Barnable, P.; Rodriguez, A.; Lai, M.; Wesenberg, A.; Ugaonkar, S.; Levendosky, K.; et al. Griffithsin carrageenan fast dissolving inserts prevent SHIV HSV-2 and HPV infections in vivo. Nat. Commun. 2018, 9, 3881. [Google Scholar] [CrossRef] [PubMed]
- Emau, P.; Tian, B.; O’Keefe, B.R.; Mori, T.; McMahon, J.B.; Palmer, K.E.; Jiang, Y.; Bekele, G.; Tsai, C.C. Griffithsin, a potent HIV entry inhibitor, is an excellent candidate for anti-HIV microbicide. J. Med. Primatol. 2007, 36, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Ferir, G.; Palmer, K.E.; Schols, D. Synergistic activity profile of griffithsin in combination with tenofovir, maraviroc and enfuvirtide against HIV-1 clade C. Virology 2011, 417, 253–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoorelbeke, B.; Xue, J.; LiWang, P.J.; Balzarini, J. Role of the carbohydrate-binding sites of griffithsin in the prevention of DC-SIGN-mediated capture and transmission of HIV-1. PLoS ONE 2013, 8, e64132. [Google Scholar] [CrossRef] [Green Version]
- Ishag, H.Z.; Li, C.; Huang, L.; Sun, M.X.; Wang, F.; Ni, B.; Malik, T.; Chen, P.Y.; Mao, X. Griffithsin inhibits Japanese encephalitis virus infection in vitro and in vivo. Arch. Virol. 2013, 158, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Ishag, H.Z.; Li, C.; Wang, F.; Mao, X. Griffithsin binds to the glycosylated proteins (E and prM) of Japanese encephalitis virus and inhibit its infection. Virus Res. 2016, 215, 50–54. [Google Scholar] [CrossRef]
- Levendosky, K.; Mizenina, O.; Martinelli, E.; Jean-Pierre, N.; Kizima, L.; Rodriguez, A.; Kleinbeck, K.; Bonnaire, T.; Robbiani, M.; Zydowsky, T.M.; et al. Griffithsin and carrageenan combination to target herpes simplex virus 2 and human papillomavirus. Antimicrob. Agents Chemother. 2015, 59, 7290–7298. [Google Scholar] [CrossRef] [Green Version]
- Meuleman, P.; Albecka, A.; Belouzard, S.; Vercauteren, K.; Verhoye, L.; Wychowski, C.; Leroux-Roels, G.; Palmer, K.E.; Dubuisson, J. Griffithsin has antiviral activity against hepatitis C virus. Antimicrob. Agents Chemother. 2011, 55, 5159–5167. [Google Scholar] [CrossRef] [Green Version]
- Micewicz, E.D.; Cole, A.L.; Jung, C.L.; Luong, H.; Phillips, M.L.; Pratikhya, P.; Sharma, S.; Waring, A.J.; Cole, A.M.; Ruchala, P. Grifonin-1: A small HIV-1 entry inhibitor derived from the algal lectin, Griffithsin. PLoS ONE 2010, 5, e14360. [Google Scholar] [CrossRef] [Green Version]
- Millet, J.K.; Whittaker, G.R. Host cell entry of Middle East respiratory syndrome coronavirus after two-step, furin-mediated activation of the spike protein. Proc. Natl. Acad. Sci. USA 2014, 111, 15214–15219. [Google Scholar] [CrossRef] [Green Version]
- Moulaei, T.; Alexandre, K.B.; Shenoy, S.R.; Meyerson, J.R.; Krumpe, L.R.; Constantine, B.; Wilson, J.; Buckheit, R.W., Jr.; McMahon, J.B.; Subramaniam, S.; et al. Griffithsin tandemers: Flexible and potent lectin inhibitors of the human immunodeficiency virus. Retrovirology 2015, 12, 6. [Google Scholar] [CrossRef] [Green Version]
- Nixon, B.; Stefanidou, M.; Mesquita, P.M.; Fakioglu, E.; Segarra, T.; Rohan, L.; Halford, W.; Palmer, K.E.; Herold, B.C. Griffithsin protects mice from genital herpes by preventing cell-to-cell spread. J. Virol. 2013, 87, 6257–6269. [Google Scholar] [CrossRef] [Green Version]
- O’Keefe, B.R.; Giomarelli, B.; Barnard, D.L.; Shenoy, S.R.; Chan, P.K.; McMahon, J.B.; Palmer, K.E.; Barnett, B.W.; Meyerholz, D.K.; Wohlford-Lenane, C.L. Broad-spectrum in vitro activity and in vivo efficacy of the antiviral protein griffithsin against emerging viruses of the family Coronaviridae. J. Virol. 2010, 84, 2511–2521. [Google Scholar] [CrossRef] [Green Version]
- O’Keefe, B.R.; Vojdani, F.; Buffa, V.; Shattock, R.J.; Montefiori, D.C.; Bakke, J.; Mirsalis, J.; d’Andrea, A.L.; Hume, S.D.; Bratcher, B.; et al. Scaleable manufacture of HIV-1 entry inhibitor griffithsin and validation of its safety and efficacy as a topical microbicide component. Proc. Natl. Acad. Sci. USA 2009, 106, 6099–6104. [Google Scholar] [CrossRef] [Green Version]
- Takebe, Y.; Saucedo, C.J.; Lund, G.; Uenishi, R.; Hase, S.; Tsuchiura, T.; Kneteman, N.; Ramessar, K.; Tyrrell, D.L.; Shirakura, M.; et al. Antiviral lectins from red and blue-green algae show potent in vitro and in vivo activity against hepatitis C virus. PLoS ONE 2013, 8, e64449. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.; Gao, Y.; Hoorelbeke, B.; Kagiampakis, I.; Zhao, B.; Demeler, B.; Balzarini, J.; Liwang, P.J. The role of individual carbohydrate-binding sites in the function of the potent anti-HIV lectin griffithsin. Mol. Pharm. 2012, 9, 2613–2625. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.; Hoorelbeke, B.; Kagiampakis, I.; Demeler, B.; Balzarini, J.; Liwang, P.J. The griffithsin dimer is required for high-potency inhibition of HIV-1: Evidence for manipulation of the structure of gp120 as part of the griffithsin dimer mechanism. Antimicrob. Agents Chemother. 2013, 57, 3976–3989. [Google Scholar] [CrossRef] [Green Version]
- Yang, E.C.; Boo, S.M. Evidence for two independent lineages of Griffithsia (Ceramiaceae, Rhodophyta) based on plastid protein-coding psaA, psbA, and rbcL gene sequences. Mol. Phylogenet. Evol. 2004, 31, 680–688. [Google Scholar] [CrossRef]
- Barrientos, L.G.; Lasala, F.; Otero, J.R.; Sanchez, A.; Delgado, R. In vitro evaluation of cyanovirin-N antiviral activity, by use of lentiviral vectors pseudotyped with filovirus envelope glycoproteins. J. Infect. Dis. 2004, 189, 1440–1443. [Google Scholar] [CrossRef] [Green Version]
- Barrientos, L.G.; O’Keefe, B.R.; Bray, M.; Sanchez, A.; Gronenborn, A.M.; Boyd, M.R. Cyanovirin-N binds to the viral surface glycoprotein, GP1,2 and inhibits infectivity of Ebola virus. Antivir. Res. 2003, 58, 47–56. [Google Scholar] [CrossRef]
- Dey, B.; Lerner, D.L.; Lusso, P.; Boyd, M.R.; Elder, J.H.; Berger, E.A. Multiple antiviral activities of cyanovirin-N: Blocking of human immunodeficiency virus type 1 gp120 interaction with CD4 and coreceptor and inhibition of diverse enveloped viruses. J. Virol. 2000, 74, 4562–4569. [Google Scholar] [CrossRef]
- O’Keefe, B.R.; Smee, D.F.; Turpin, J.A.; Saucedo, C.J.; Gustafson, K.R.; Mori, T.; Blakeslee, D.; Buckheit, R.; Boyd, M.R. Potent anti-influenza activity of cyanovirin-N and interactions with viral hemagglutinin. Antimicrob. Agents Chemother. 2003, 47, 2518–2525. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.S.; Walia, A.K.; Khattar, J.S.; Singh, D.P.; Kennedy, J.F. Cyanobacterial lectins characteristics and their role as antiviral agents. Int. J. Biol. Macromol. 2017, 102, 475–496. [Google Scholar] [CrossRef]
- Smee, D.F.; Bailey, K.W.; Wong, M.H.; O’Keefe, B.R.; Gustafson, K.R.; Mishin, V.P.; Gubareva, L.V. Treatment of influenza A (H1N1) virus infections in mice and ferrets with cyanovirin-N. Antivir. Res. 2008, 80, 266–271. [Google Scholar] [CrossRef] [Green Version]
- Smee, D.F.; Wandersee, M.K.; Checketts, M.B.; O’Keefe, B.R.; Saucedo, C.; Boyd, M.R.; Mishin, V.P.; Gubareva, L.V. Influenza A (H1N1) virus resistance to cyanovirin-N arises naturally during adaptation to mice and by passage in cell culture in the presence of the inhibitor. Antivir. Chem. Chemother. 2007, 18, 317–327. [Google Scholar] [CrossRef] [Green Version]
- van Gremberghe, I.; Leliaert, F.; Mergeay, J.; Vanormelingen, P.; Van der Gucht, K.; Debeer, A.E.; Lacerot, G.; De Meester, L.; Vyverman, W. Lack of phylogeographic structure in the freshwater cyanobacterium Microcystis aeruginosa suggests global dispersal. PLoS ONE 2011, 6, e19561. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Okuyama, S.; Hori, K. Primary structure and carbohydrate binding specificity of a potent anti-HIV lectin isolated from the filamentous cyanobacterium Oscillatoria agardhii. J. Biol. Chem. 2007, 282, 11021–11029. [Google Scholar] [CrossRef] [Green Version]
- Holanda, M.L.; Melo, V.M.; Silva, L.M.; Amorim, R.C.; Pereira, M.G.; Benevides, N.M. Differential activity of a lectin from Solieria filiformis against human pathogenic bacteria. Braz. J. Med. Biol. Res. 2005, 38, 1769–1773. [Google Scholar] [CrossRef] [Green Version]
- Hirayama, M.; Shibata, H.; Imamura, K.; Sakaguchi, T.; Hori, K. High-mannose specific lectin and its recombinants from a carrageenophyta kappaphycus alvarezii represent a potent anti-HIV activity through high-affinity binding to the viral envelope glycoprotein gp120. Mar. Biotechnol. 2016, 18, 144–160. [Google Scholar] [CrossRef]
- Sato, Y.; Morimoto, K.; Hirayama, M.; Hori, K. High mannose-specific lectin (KAA-2) from the red alga Kappaphycus alvarezii potently inhibits influenza virus infection in a strain-independent manner. Biochem. Biophys. Res. Commun. 2011, 405, 291–296. [Google Scholar] [CrossRef]
- Sato, Y.; Morimoto, K.; Kubo, T.; Sakaguchi, T.; Nishizono, A.; Hirayama, M.; Hori, K. Entry inhibition of influenza viruses with high mannose binding lectin ESA-2 from the red alga eucheuma serra through the recognition of viral hemagglutinin. Mar. Drugs 2015, 13, 3454–3465. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Hirayama, M.; Morimoto, K.; Yamamoto, N.; Okuyama, S.; Hori, K. High mannose-binding lectin with preference for the cluster of alpha1-2-mannose from the green alga Boodlea coacta is a potent entry inhibitor of HIV-1 and influenza viruses. J. Biol. Chem. 2011, 286, 19446–19458. [Google Scholar] [CrossRef] [Green Version]
- Mu, J.; Hirayama, M.; Sato, Y.; Morimoto, K.; Hori, K. A novel high-mannose specific lectin from the green alga halimeda renschii exhibits a potent anti-influenza virus activity through high-affinity binding to the viral hemagglutinin. Mar. Drugs 2017, 15, 255. [Google Scholar] [CrossRef] [Green Version]
- Garrison, A.R.; Giomarelli, B.G.; Lear-Rooney, C.M.; Saucedo, C.J.; Yellayi, S.; Krumpe, L.R.; Rose, M.; Paragas, J.; Bray, M.; Olinger, G.G., Jr.; et al. The cyanobacterial lectin scytovirin displays potent in vitro and in vivo activity against Zaire Ebola virus. Antivir. Res. 2014, 112, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Alexandre, K.B.; Gray, E.S.; Lambson, B.E.; Moore, P.L.; Choge, I.A.; Mlisana, K.; Karim, S.S.; McMahon, J.; O’Keefe, B.; Chikwamba, R.; et al. Mannose-rich glycosylation patterns on HIV-1 subtype C gp120 and sensitivity to the lectins, Griffithsin, Cyanovirin-N and Scytovirin. Virology 2010, 402, 187–196. [Google Scholar] [CrossRef] [Green Version]
- Balzarini, J. Targeting the glycans of gp120: A novel approach aimed at the Achilles heel of HIV. Lancet Infect. Dis. 2005, 5, 726–731. [Google Scholar] [CrossRef]
- Rudd, P.M.; Elliott, T.; Cresswell, P.; Wilson, I.A.; Dwek, R.A. Glycosylation and the immune system. Science 2001, 291, 2370–2376. [Google Scholar] [CrossRef]
- Daniels, C.N.; Saunders, K.O. Chapter 2: Antibody responses to the HIV-1 envelope high mannose patch. Adv. Immunol. 2019, 143, 11–73. [Google Scholar]
- Coss, K.P.; Vasiljevic, S.; Pritchard, L.K.; Krumm, S.A.; Glaze, M.; Madzorera, S.; Moore, P.L.; Crispin, M.; Doores, K.J. HIV-1 glycan density drives the persistence of the mannose patch within an infected individual. J. Virol. 2016, 90, 11132–11144. [Google Scholar] [CrossRef] [Green Version]
- Bonsignori, M.; Hwang, K.K.; Chen, X.; Tsao, C.Y.; Morris, L.; Gray, E.; Marshall, D.J.; Crump, J.A.; Kapiga, S.H.; Sam, N.E.; et al. Analysis of a clonal lineage of HIV-1 envelope V2/V3 conformational epitope-specific broadly neutralizing antibodies and their inferred unmutated common ancestors. J. Virol. 2011, 85, 9998–10009. [Google Scholar] [CrossRef] [Green Version]
- Doria-Rose, N.A.; Bhiman, J.N.; Roark, R.S.; Schramm, C.A.; Gorman, J.; Chuang, G.Y.; Pancera, M.; Cale, E.M.; Ernandes, M.J.; Louder, M.K.; et al. New member of the V1V2-directed CAP256-VRC26 lineage that shows increased breadth and exceptional potency. J. Virol. 2016, 90, 76–91. [Google Scholar] [CrossRef] [Green Version]
- Falkowska, E.; Le, K.M.; Ramos, A.; Doores, K.J.; Lee, J.H.; Blattner, C.; Ramirez, A.; Derking, R.; van Gils, M.J.; Liang, C.H.; et al. Broadly neutralizing HIV antibodies define a glycan-dependent epitope on the prefusion conformation of gp41 on cleaved envelope trimers. Immunity 2014, 40, 657–668. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Kang, B.H.; Pancera, M.; Lee, J.H.; Tong, T.; Feng, Y.; Imamichi, H.; Georgiev, I.S.; Chuang, G.Y.; Druz, A.; et al. Broad and potent HIV-1 neutralization by a human antibody that binds the gp41-gp120 interface. Nature 2014, 515, 138–142. [Google Scholar] [CrossRef]
- Kong, L.; Lee, J.H.; Doores, K.J.; Murin, C.D.; Julien, J.P.; McBride, R.; Liu, Y.; Marozsan, A.; Cupo, A.; Klasse, P.J.; et al. Supersite of immune vulnerability on the glycosylated face of HIV-1 envelope glycoprotein gp120. Nat. Struct. Mol. Biol. 2013, 20, 796–803. [Google Scholar] [CrossRef] [Green Version]
- Pejchal, R.; Doores, K.J.; Walker, L.M.; Khayat, R.; Huang, P.S.; Wang, S.K.; Stanfield, R.L.; Julien, J.P.; Ramos, A.; Crispin, M.; et al. A potent and broad neutralizing antibody recognizes and penetrates the HIV glycan shield. Science 2011, 334, 1097–1103. [Google Scholar] [CrossRef] [Green Version]
- Scharf, L.; Scheid, J.F.; Lee, J.H.; West, A.P., Jr.; Chen, C.; Gao, H.; Gnanapragasam, P.N.; Mares, R.; Seaman, M.S.; Ward, A.B.; et al. Antibody 8ANC195 reveals a site of broad vulnerability on the HIV-1 envelope spike. Cell Rep. 2014, 7, 785–795. [Google Scholar] [CrossRef] [Green Version]
- Schoning, G.; Plonait, H. Metaphylaxis and therapy of the MMA syndrome of sows with Baytril. Dtsch. Tierarztl. Wochenschr. 1990, 97, 5–10. [Google Scholar]
- Walker, L.M.; Phogat, S.K.; Chan-Hui, P.Y.; Wagner, D.; Phung, P.; Goss, J.L.; Wrin, T.; Simek, M.D.; Fling, S.; Mitcham, J.L.; et al. Broad and potent neutralizing antibodies from an African donor reveal a new HIV-1 vaccine target. Science 2009, 326, 285–289. [Google Scholar] [CrossRef] [Green Version]
- Balzarini, J.; Van Damme, L. Microbicide drug candidates to prevent HIV infection. Lancet 2007, 369, 787–797. [Google Scholar] [CrossRef]
- Huang, X.; Jin, W.; Griffin, G.E.; Shattock, R.J.; Hu, Q. Removal of two high-mannose N-linked glycans on gp120 renders human immunodeficiency virus 1 largely resistant to the carbohydrate-binding agent griffithsin. J. Gen. Virol. 2011, 92, 2367–2373. [Google Scholar] [CrossRef]
- Alexandre, K.B.; Gray, E.S.; Mufhandu, H.; McMahon, J.B.; Chakauya, E.; O’Keefe, B.R.; Chikwamba, R.; Morris, L. The lectins griffithsin, cyanovirin-N and scytovirin inhibit HIV-1 binding to the DC-SIGN receptor and transfer to CD4(+) cells. Virology 2012, 423, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Balzarini, J.; Van Herrewege, Y.; Vermeire, K.; Vanham, G.; Schols, D. Carbohydrate-binding agents efficiently prevent dendritic cell-specific intercellular adhesion molecule-3-grabbing nonintegrin (DC-SIGN)-directed HIV-1 transmission to T lymphocytes. Mol. Pharmacol. 2007, 71, 3–11. [Google Scholar] [CrossRef]
- Hu, B.; Du, T.; Li, C.; Luo, S.; Liu, Y.; Huang, X.; Hu, Q. Sensitivity of transmitted and founder human immunodeficiency virus type 1 envelopes to carbohydrate-binding agents griffithsin, cyanovirin-N and Galanthus nivalis agglutinin. J. Gen. Virol. 2015, 96, 3660–3666. [Google Scholar] [CrossRef]
- Alexandre, K.B.; Moore, P.L.; Nonyane, M.; Gray, E.S.; Ranchobe, N.; Chakauya, E.; McMahon, J.B.; O’Keefe, B.R.; Chikwamba, R.; Morris, L. Mechanisms of HIV-1 subtype C resistance to GRFT, CV-N and SVN. Virology 2013, 446, 66–76. [Google Scholar] [CrossRef] [Green Version]
- Huskens, D.; Ferir, G.; Vermeire, K.; Kehr, J.C.; Balzarini, J.; Dittmann, E.; Schols, D. Microvirin, a novel alpha(1,2)-mannose-specific lectin isolated from Microcystis aeruginosa, has anti-HIV-1 activity comparable with that of cyanovirin-N but a much higher safety profile. J. Biol. Chem. 2010, 285, 24845–24854. [Google Scholar] [CrossRef] [Green Version]
- Ferir, G.; Huskens, D.; Palmer, K.E.; Boudreaux, D.M.; Swanson, M.D.; Markovitz, D.M.; Balzarini, J.; Schols, D. Combinations of griffithsin with other carbohydrate-binding agents demonstrate superior activity against HIV Type 1, HIV Type 2, and selected carbohydrate-binding agent-resistant HIV Type 1 strains. AIDS Res. Hum. Retrovir. 2012, 28, 1513–1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellett, P.E.; Roizman, B. Herpesviridae. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott-Raven Publishers: Philadelphia, PA, USA, 2013; pp. 1802–1822. [Google Scholar]
- Chiang, H.Y.; Cohen, G.H.; Eisenberg, R.J. Identification of functional regions of herpes simplex virus glycoprotein gD by using linker-insertion mutagenesis. J. Virol. 1994, 68, 2529–2543. [Google Scholar] [CrossRef] [Green Version]
- Leumi, S.; El Kassas, M.; Zhong, J. Hepatitis C virus genotype 4: A poorly characterized endemic genotype. J. Med. Virol. 2021, 93, 6079–6088. [Google Scholar] [CrossRef]
- Moradpour, D.; Penin, F.; Rice, C.M. Replication of hepatitis C virus. Nat. Rev. Microbiol. 2007, 5, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Bukh, J.; Kuiken, C.; Muerhoff, A.S.; Rice, C.M.; Stapleton, J.T.; Simmonds, P. Expanded classification of hepatitis C virus into 7 genotypes and 67 subtypes: Updated criteria and genotype assignment web resource. Hepatology 2014, 59, 318–327. [Google Scholar] [CrossRef] [Green Version]
- Structural biology of the hepatitis C virus proteins. Drug Discov. Today Technol. 2012, 9, e175–e226.
- Dubuisson, J. Hepatitis C virus proteins. World J. Gastroenterol. 2007, 13, 2406–2415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.W.; Chang, K.M. Hepatitis C virus: Virology and life cycle. Clin. Mol. Hepatol. 2013, 19, 17–25. [Google Scholar] [CrossRef]
- Krey, T.; d’Alayer, J.; Kikuti, C.M.; Saulnier, A.; Damier-Piolle, L.; Petitpas, I.; Johansson, D.X.; Tawar, R.G.; Baron, B.; Robert, B.; et al. The disulfide bonds in glycoprotein E2 of hepatitis C virus reveal the tertiary organization of the molecule. PLoS Pathog. 2010, 6, e1000762. [Google Scholar] [CrossRef]
- Benedicto, I.; Gondar, V.; Molina-Jimenez, F.; Garcia-Buey, L.; Lopez-Cabrera, M.; Gastaminza, P.; Majano, P.L. Clathrin mediates infectious hepatitis C virus particle egress. J. Virol. 2015, 89, 4180–4190. [Google Scholar] [CrossRef] [Green Version]
- Blanchard, E.; Belouzard, S.; Goueslain, L.; Wakita, T.; Dubuisson, J.; Wychowski, C.; Rouille, Y. Hepatitis C virus entry depends on clathrin-mediated endocytosis. J. Virol. 2006, 80, 6964–6972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindenbach, B.D.; Rice, C.M. The ins and outs of hepatitis C virus entry and assembly. Nat. Rev. Microbiol. 2013, 11, 688–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goffard, A.; Callens, N.; Bartosch, B.; Wychowski, C.; Cosset, F.L.; Montpellier, C.; Dubuisson, J. Role of N-linked glycans in the functions of hepatitis C virus envelope glycoproteins. J. Virol. 2005, 79, 8400–8409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieyres, G.; Thomas, X.; Descamps, V.; Duverlie, G.; Patel, A.H.; Dubuisson, J. Characterization of the envelope glycoproteins associated with infectious hepatitis C virus. J. Virol. 2010, 84, 10159–10168. [Google Scholar] [CrossRef] [Green Version]
- Helle, F.; Wychowski, C.; Vu-Dac, N.; Gustafson, K.R.; Voisset, C.; Dubuisson, J. Cyanovirin-N inhibits hepatitis C virus entry by binding to envelope protein glycans. J. Biol. Chem. 2006, 281, 25177–25183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kachko, A.; Loesgen, S.; Shahzad-Ul-Hussan, S.; Tan, W.; Zubkova, I.; Takeda, K.; Wells, F.; Rubin, S.; Bewley, C.A.; Major, M.E. Inhibition of hepatitis C virus by the cyanobacterial protein Microcystis viridis lectin: Mechanistic differences between the high-mannose specific lectins MVL, CV-N, and GNA. Mol. Pharm. 2013, 10, 4590–4602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C. Griffithsin, a highly potent broad-spectrum antiviral lectin from red algae: From discovery to clinical application. Mar. Drugs 2019, 17, 567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lusvarghi, S.; Bewley, C.A. Griffithsin: An antiviral lectin with outstanding therapeutic potential. Viruses 2016, 8, 296. [Google Scholar] [CrossRef] [PubMed]
- Nuwarda, R.F.; Alharbi, A.A.; Kayser, V. An overview of influenza viruses and vaccines. Vaccines 2021, 9, 1032. [Google Scholar] [CrossRef]
- Saunders-Hastings, P.R.; Krewski, D. Reviewing the history of pandemic influenza: Understanding patterns of emergence and transmission. Pathogens 2016, 5, 66. [Google Scholar] [CrossRef] [Green Version]
- Nayak, D.P.; Balogun, R.A.; Yamada, H.; Zhou, Z.H.; Barman, S. Influenza virus morphogenesis and budding. Virus Res. 2009, 143, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Moules, V.; Terrier, O.; Yver, M.; Riteau, B.; Moriscot, C.; Ferraris, O.; Julien, T.; Giudice, E.; Rolland, J.P.; Erny, A.; et al. Importance of viral genomic composition in modulating glycoprotein content on the surface of influenza virus particles. Virology 2011, 414, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Gamblin, S.J.; Skehel, J.J. Influenza hemagglutinin and neuraminidase membrane glycoproteins. J. Biol. Chem. 2010, 285, 28403–28409. [Google Scholar] [CrossRef] [Green Version]
- Skehel, J.J.; Wiley, D.C. Receptor binding and membrane fusion in virus entry: The influenza hemagglutinin. Annu. Rev. Biochem. 2000, 69, 531–569. [Google Scholar] [CrossRef] [PubMed]
- Gaymard, A.; Le Briand, N.; Frobert, E.; Lina, B.; Escuret, V. Functional balance between neuraminidase and haemagglutinin in influenza viruses. Clin. Microbiol. Infect. 2016, 22, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Reading, P.C.; Tate, M.D.; Pickett, D.L.; Brooks, A.G. Glycosylation as a target for recognition of influenza viruses by the innate immune system. Adv. Exp. Med. Biol. 2007, 598, 279–292. [Google Scholar]
- Wu, C.; Chen, W.; Chen, J.; Han, B.; Peng, Z.; Ge, F.; Wei, B.; Liu, M.; Zhang, M.; Qian, C.; et al. Preparation of monoPEGylated Cyanovirin-N’s derivative and its anti-influenza A virus bioactivity in vitro and in vivo. J. Biochem. 2015, 157, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Baseler, L.; Chertow, D.S.; Johnson, K.M.; Feldmann, H.; Morens, D.M. The pathogenesis of Ebola virus disease. Annu. Rev. Pathol. 2017, 12, 387–418. [Google Scholar] [CrossRef] [PubMed]
- Furuyama, W.; Marzi, A. Ebola virus: Pathogenesis and countermeasure development. Annu. Rev. Virol. 2019, 6, 435–458. [Google Scholar] [CrossRef]
- Grifoni, A.; Lo Presti, A.; Giovanetti, M.; Montesano, C.; Amicosante, M.; Colizzi, V.; Lai, A.; Zehender, G.; Cella, E.; Angeletti, S.; et al. Genetic diversity in Ebola virus: Phylogenetic and in silico structural studies of Ebola viral proteins. Asian Pac. J. Trop. Med. 2016, 9, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Stahelin, R.V. Membrane binding and bending in Ebola VP40 assembly and egress. Front. Microbiol. 2014, 5, 300. [Google Scholar] [PubMed] [Green Version]
- Jeffers, S.A.; Sanders, D.A.; Sanchez, A. Covalent modifications of the ebola virus glycoprotein. J. Virol. 2002, 76, 12463–12472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volchkov, V.E.; Feldmann, H.; Volchkova, V.A.; Klenk, H.D. Processing of the Ebola virus glycoprotein by the proprotein convertase furin. Proc. Natl. Acad. Sci. USA 1998, 95, 5762–5767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.E.; Fusco, M.L.; Hessell, A.J.; Oswald, W.B.; Burton, D.R.; Saphire, E.O. Structure of the Ebola virus glycoprotein bound to an antibody from a human survivor. Nature 2008, 454, 177–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.E.; Saphire, E.O. Ebolavirus glycoprotein structure and mechanism of entry. Future Virol. 2009, 4, 621–635. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, G.; Harvey, D.J.; Stroeher, U.; Feldmann, F.; Feldmann, H.; Wahl-Jensen, V.; Royle, L.; Dwek, R.A.; Rudd, P.M. Identification of N-glycans from Ebola virus glycoproteins by matrix-assisted laser desorption/ionisation time-of-flight and negative ion electrospray tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 571–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barre, A.; Van Damme, E.J.M.; Simplicien, M.; Le Poder, S.; Klonjkowski, B.; Benoist, H.; Peyrade, D.; Rouge, P. Man-specific lectins from plants, fungi, algae and cyanobacteria, as potential blockers for SARS-CoV, MERS-CoV and SARS-CoV-2 (COVID-19) coronaviruses: Biomedical perspectives. Cells 2021, 10, 1619. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Xu, W.; Gu, C.; Cai, X.; Qu, D.; Lu, L.; Xie, Y.; Jiang, S. Griffithsin with a broad-spectrum antiviral activity by binding glycans in viral glycoprotein exhibits strong synergistic effect in combination with a pan-coronavirus fusion inhibitor targeting SARS-CoV-2 Spike S2 subunit. Virol. Sin. 2020, 35, 857–860. [Google Scholar] [CrossRef] [PubMed]
- Millet, J.K.; Seron, K.; Labitt, R.N.; Danneels, A.; Palmer, K.E.; Whittaker, G.R.; Dubuisson, J.; Belouzard, S. Middle East respiratory syndrome coronavirus infection is inhibited by griffithsin. Antivir. Res. 2016, 133, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bhowmick, S.; Mazumdar, A.; Moulick, A.; Adam, V. Algal metabolites: An inevitable substitute for antibiotics. Biotechnol. Adv. 2020, 43, 107571. [Google Scholar] [CrossRef]
- Breitenbach Barroso Coelho, L.C.; Marcelino Dos Santos Silva, P.; Felix de Oliveira, W.; de Moura, M.C.; Viana Pontual, E.; Soares Gomes, F.; Guedes Paiva, P.M.; Napoleao, T.H.; Dos Santos Correia, M.T. Lectins as antimicrobial agents. J. Appl. Microbiol. 2018, 125, 1238–1252. [Google Scholar] [CrossRef] [Green Version]
- Liao, W.R.; Lin, J.Y.; Shieh, W.Y.; Jeng, W.L.; Huang, R. Antibiotic activity of lectins from marine algae against marine vibrios. J. Ind. Microbiol. Biotechnol. 2003, 30, 433–439. [Google Scholar] [CrossRef]
- Hung, L.D.; Hirayama, M.; Ly, B.M.; Hori, K. Purification, primary structure, and biological activity of the high-mannose N-glycan-specific lectin from cultivated Eucheuma denticulatum. J. Appl. Phycol. 2015, 27, 1657–1669. [Google Scholar] [CrossRef] [PubMed]
- Gilljam, H.; Thiringer, G. The right to a smoke-free occupational environment is a reasonable and natural demand. Lakartidningen 1992, 89, 1977–1978. [Google Scholar] [PubMed]
- Vasconcelos, M.A.; Arruda, F.V.; Carneiro, V.A.; Silva, H.C.; Nascimento, K.S.; Sampaio, A.H.; Cavada, B.; Teixeira, E.H.; Henriques, M.; Pereira, M.O. Effect of algae and plant lectins on planktonic growth and biofilm formation in clinically relevant bacteria and yeasts. Biomed. Res. Int. 2014, 2014, 365272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Felicio, R.; de Albuquerque, S.; Young, M.C.; Yokoya, N.S.; Debonsi, H.M. Trypanocidal, leishmanicidal and antifungal potential from marine red alga Bostrychia tenella J. Agardh (Rhodomelaceae, ceramiales). J. Pharm. Biomed. Anal. 2010, 52, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Bianco, E.M.; de Oliveira, S.Q.; Rigotto, C.; Tonini, M.L.; da Rosa Guimaraes, T.; Bittencourt, F.; Gouvea, L.P.; Aresi, C.; de Almeida, M.T.; Moritz, M.I.; et al. Anti-infective potential of marine invertebrates and seaweeds from the Brazilian coast. Molecules 2013, 18, 5761–5778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Alarcon, D.; Blanco-Labra, A.; Garcia-Gasca, T. Expression of lectins in heterologous systems. Int. J. Mol. Sci. 2018, 19, 616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, A.; Jiang, L.; Kittleson, G.A.; Steadman, K.D.; Nandi, S.; Fuqua, J.L.; Palmer, K.E.; Tuse, D.; McDonald, K.A. Technoeconomic modeling of plant-based griffithsin manufacturing. Front. Bioeng. Biotechnol. 2018, 6, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habibi, P.; Soccol, C.R.; O’Keefe, B.R.; Krumpe, L.R.H.; Wilson, J.; de Macedo, L.L.P.; Faheem, M.; Dos Santos, V.O.; Prado, G.S.; Botelho, M.A.; et al. Gene-silencing suppressors for high-level production of the HIV-1 entry inhibitor griffithsin in Nicotiana benthamiana. Process. Biochem. 2018, 70, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Sexton, A.; Drake, P.M.; Mahmood, N.; Harman, S.J.; Shattock, R.J.; Ma, J.K. Transgenic plant production of Cyanovirin-N, an HIV microbicide. FASEB J. 2006, 20, 356–358. [Google Scholar] [CrossRef]
- Mackie, D.M.; Cohen, R.J.; Glick, A.J. Soliton contributions to the third-order susceptibility of polyacetylene. Phys. Rev. B Condens. Matter 1989, 39, 3442–3444. [Google Scholar] [CrossRef]
- Vamvaka, E.; Arcalis, E.; Ramessar, K.; Evans, A.; O’Keefe, B.R.; Shattock, R.J.; Medina, V.; Stoger, E.; Christou, P.; Capell, T. Rice endosperm is cost-effective for the production of recombinant griffithsin with potent activity against HIV. Plant Biotechnol. J. 2016, 14, 1427–1437. [Google Scholar] [CrossRef] [Green Version]
- Giomarelli, B.; Schumacher, K.M.; Taylor, T.E.; Sowder, R.C., 2nd; Hartley, J.L.; McMahon, J.B.; Mori, T. Recombinant production of anti-HIV protein, griffithsin, by auto-induction in a fermentor culture. Protein Expr. Purif. 2006, 47, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Decker, J.S.; Menacho-Melgar, R.; Lynch, M.D. Low-cost, large-scale production of the anti-viral lectin griffithsin. Front. Bioeng. Biotechnol. 2020, 8, 1020. [Google Scholar] [CrossRef] [PubMed]
- Petrova, M.I.; van den Broek, M.F.L.; Spacova, I.; Verhoeven, T.L.A.; Balzarini, J.; Vanderleyden, J.; Schols, D.; Lebeer, S. Engineering lactobacillus rhamnosus GG and GR-1 to express HIV-inhibiting griffithsin. Int. J. Antimicrob. Agents 2018, 52, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lagenaur, L.A.; Simpson, D.A.; Essenmacher, K.P.; Frazier-Parker, C.L.; Liu, Y.; Tsai, D.; Rao, S.S.; Hamer, D.H.; Parks, T.P.; et al. Engineered vaginal lactobacillus strain for mucosal delivery of the human immunodeficiency virus inhibitor cyanovirin-N. Antimicrob. Agents Chemother. 2006, 50, 3250–3259. [Google Scholar] [CrossRef] [Green Version]
- Pusch, O.; Boden, D.; Hannify, S.; Lee, F.; Tucker, L.D.; Boyd, M.R.; Wells, J.M.; Ramratnam, B. Bioengineering lactic acid bacteria to secrete the HIV-1 virucide cyanovirin. J. Acquir. Immune Defic. Syndr. 2005, 40, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.S.; Xu, Q.; Fichorova, R.N. Homeostatic properties of Lactobacillus jensenii engineered as a live vaginal anti-HIV microbicide. BMC Microbiol. 2013, 13, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janahi, E.M.A.; Haque, S.; Akhter, N.; Wahid, M.; Jawed, A.; Mandal, R.K.; Lohani, M.; Areeshi, M.Y.; Almalki, S.; Das, S.; et al. Bioengineered intravaginal isolate of Lactobacillus plantarum expresses algal lectin scytovirin demonstrating anti-HIV-1 activity. Microb. Pathog. 2018, 122, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Romero, J.A.; Teleshova, N.; Zydowsky, T.M.; Robbiani, M. Preclinical assessments of vaginal microbicide candidate safety and efficacy. Adv. Drug Deliv. Rev. 2015, 92, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Lagenaur, L.A.; Sanders-Beer, B.E.; Brichacek, B.; Pal, R.; Liu, X.; Liu, Y.; Yu, R.; Venzon, D.; Lee, P.P.; Hamer, D.H. Prevention of vaginal SHIV transmission in macaques by a live recombinant Lactobacillus. Mucosal. Immunol. 2011, 4, 648–657. [Google Scholar] [CrossRef] [Green Version]
- Buffa, V.; Stieh, D.; Mamhood, N.; Hu, Q.; Fletcher, P.; Shattock, R.J. Cyanovirin-N potently inhibits human immunodeficiency virus type 1 infection in cellular and cervical explant models. J. Gen. Virol. 2009, 90, 234–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huskens, D.; Vermeire, K.; Vandemeulebroucke, E.; Balzarini, J.; Schols, D. Safety concerns for the potential use of cyanovirin-N as a microbicidal anti-HIV agent. Int. J. Biochem. Cell. Biol. 2008, 40, 2802–2814. [Google Scholar] [CrossRef] [PubMed]
- Balzarini, J.; Van Laethem, K.; Peumans, W.J.; Van Damme, E.J.; Bolmstedt, A.; Gago, F.; Schols, D. Mutational pathways, resistance profile, and side effects of cyanovirin relative to human immunodeficiency virus type 1 strains with N-glycan deletions in their gp120 envelopes. J. Virol. 2006, 80, 8411–8421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zappe, H.; Snell, M.E.; Bossard, M.J. PEGylation of cyanovirin-N, an entry inhibitor of HIV. Adv. Drug Deliv. Rev. 2008, 60, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Shahzad-ul-Hussan, S.; Gustchina, E.; Ghirlando, R.; Clore, G.M.; Bewley, C.A. Solution structure of the monovalent lectin microvirin in complex with Man(alpha) (1–2) Man provides a basis for anti-HIV activity with low toxicity. J. Biol Chem. 2011, 286, 20788–20796. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, X. Recombinant microcystis viridis lectin as a potential anticancer agent. Pharmazie 2010, 65, 922–923. [Google Scholar]
- Sato, Y.; Murakami, M.; Miyazawa, K.; Hori, K. Purification and characterization of a novel lectin from a freshwater cyanobacterium, Oscillatoria agardhii. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2000, 125, 169–177. [Google Scholar] [CrossRef]
- Kouokam, J.C.; Huskens, D.; Schols, D.; Johannemann, A.; Riedell, S.K.; Walter, W.; Walker, J.M.; Matoba, N.; O’Keefe, B.R.; Palmer, K.E. Investigation of griffithsin’s interactions with human cells confirms its outstanding safety and efficacy profile as a microbicide candidate. PLoS ONE 2011, 6, e22635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, C.; Kouokam, J.C.; Lasnik, A.B.; Foreman, O.; Cambon, A.; Brock, G.; Montefiori, D.C.; Vojdani, F.; McCormick, A.A.; O’Keefe, B.R.; et al. Activity of and effect of subcutaneous treatment with the broad-spectrum antiviral lectin griffithsin in two laboratory rodent models. Antimicrob. Agents Chemother. 2014, 58, 120–127. [Google Scholar] [CrossRef] [Green Version]
- US National Library of Medicine. Griffithsin-Based Rectal Microbicide for PREvention of Viral ENTry (PREVENT) Clinical Trials Database. 2020. Available online: https://clinicaltrials.gov/ct2/show/NCT04032717 (accessed on 31 October 2021).
Figure 1.
Three-Dimensional Structure of Griffithsin (GRFT)-Mannose Complex (PDB ID 2GUD at 0.94 Å resolution). Left-Panel: GRFT domain-swapped dimer. Six carbohydrate (mannose)-binding sites are shown; three for each monomer in yellow and purple, respectively. Right-Panel: Magnification of the GRFT six mannose-binding sites shown in the presence (left) and absence (right) of the β-strands cartoon representation. GRFT-Mannoses main interactions via hydrogen bonds are shown as yellow dashed lines. The three GRFT mannose-binding sites together form an almost perfect equilateral triangle.
Figure 1.
Three-Dimensional Structure of Griffithsin (GRFT)-Mannose Complex (PDB ID 2GUD at 0.94 Å resolution). Left-Panel: GRFT domain-swapped dimer. Six carbohydrate (mannose)-binding sites are shown; three for each monomer in yellow and purple, respectively. Right-Panel: Magnification of the GRFT six mannose-binding sites shown in the presence (left) and absence (right) of the β-strands cartoon representation. GRFT-Mannoses main interactions via hydrogen bonds are shown as yellow dashed lines. The three GRFT mannose-binding sites together form an almost perfect equilateral triangle.

Figure 2.
Strategies to produce and purify GRFT in heterologous systems. Created with BioRender.com.
Figure 2.
Strategies to produce and purify GRFT in heterologous systems. Created with BioRender.com.


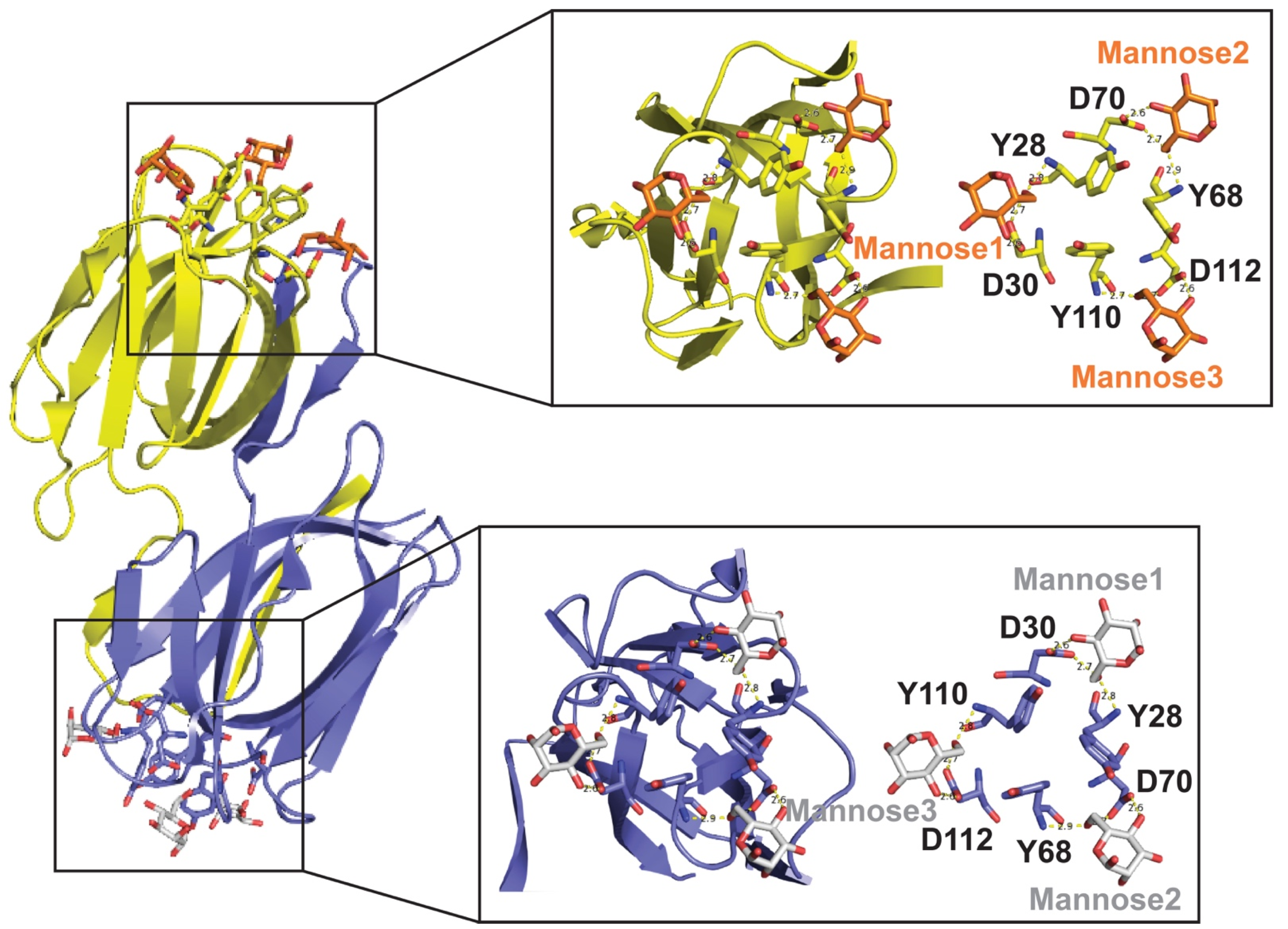{kind=link}
{kind=link}
Table 1.
Marine and freshwater lectins produced by algae and cyanobacteria.
| Lectin | Lectin Family | Carbohydrate-Binding Specificity | Marine Species That Produce Each Lectin | Antimicrobial Properties | References |
|---|---|---|---|---|---|
| GRFT | JRL | High-mannose glycans HIV-1 gp120 KD j 80 pM | Produced by Griffithsia sp. Griffithsia is a genus of red algae that includes about 27 species worldwide, most of which occur intertidally along temperate to tropical oceans in Europe, South America, and Africa. | HIV a HSV b HCV c SARS-CoV-1 and 2 d MERS CoV e EBOV f JEV g HPV h Trichomonas vaginalis | [2,7,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35] |
| CV-N | CVNHs | High-mannose glycans HIV-1 gp120 KD 6–45 nM | Produced by Nostoc ellipsosporum. Nostoc is a genus of oxygenic photosynthetic cyanobacteria that are widely distributed in terrestrial and aquatic habitats. Nostoc can resist desiccation, freezing, and thawing; and can fix atmospheric N2. | HIV HCV Influenza virus Rhinoviruses SARS-CoV-2 EBOV Measles virus HHV-6 i Trichomonas vaginalis | [17,36,37,38,39,40,41,42] |
| MVL | CVNHs | High-mannose glycans HIV-1 gp120 KD 70–100 nM | MVL and MVN are produced by Microcystis viridis and Microcystis aeruginosa, respectively. Microcystis is a genus of freshwater cyanobacteria. They are planktonic and thrive in warm, stagnant waters worldwide. | HIV-1 HCV | [43] |
| Microvirin (MVN) | CVNHs | High-mannose glycans HIV-1 gp120 KD | |||
| OAA | OAAH | High-mannose glycans HIV-1 gp120 KD 0.6 nM | Produced by Oscillatoria agardhii. Oscillatoria is a genus of filamentous cyanobacteria. The genus name refers to the oscillation in the organism’s movement. They are found in freshwater. | HIV-1 | [44] |
| SfL | OAAH | High-mannose glycans | Produced by Solieria filiformis. S. filiformis is a red alga found in subtropical warm waters along the coast in the North Atlantic Ocean, Gulf of Mexico, Colombia, Brazil, West Africa, Mediterranean, Israel, and Arabian Gulf. | Serratia marcescens Salmonella typhi Klebsiella pneumoniae Enterobacter aerogenes Proteus sp. Pseudomonas aeruginosa | [45] |
| KAA-2 | OAAH | High-mannose glycans | Produced by Kappaphycus alvarezii. K. alvarezii is a red alga that occurs naturally in the south of the Philippines and is also cultivated in the South Pacific. K. alvarezii grows in a wide variety of environments. | HIV-1 Influenza virus | [40,46,47,48] |
| BCA | GNA-like | High-mannose glycans HIV-1 gp120 KD 2.7 nM | Produced by Boodlea coacta. B. coacta is a green alga widespread throughout the tropics and can be seasonally dominant on some Indo-West Pacific reef-flats. | HIV-1 Influenza virus | [12,49] |
| HRL40 | Unknown | High-mannose glycans | Produced by Halimeda renschii. H. renschii is a segmented-marine green alga that occurs naturally in the Indo-Pacific region and the Atlantic Ocean. It is a major contributor to marine sediments in the tropics and subtropics. | Influenza virus | [50] |
| SVN | Unknown | High-mannose glycans | Produced by Scytonema varium. Scytonema includes cyanobacteria generally considered to be cosmopolitan. S. varium is a freshwater or terrestrial species. | HIV HCV SARS-CoV-1 EBOV | [51] |
a HIV: human immunodeficiency virus; b HSV: herpes simplex virus; c HCV: hepatitis C virus; d SARS-CoV-1 and 2: severe respiratory syndrome coronavirus 1 and 2; e MERS-CoV: Middle East respiratory syndrome coronavirus; f EBOV: Ebola virus; g JEV: Japanese encephalitis virus; h HPV: human papillomavirus; I HHV-6: human herpes virus 6. j KD = dissociation constant after binding to HIV-1 gp120.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fernández Romero, J.A.; Paglini, M.G.; Priano, C.; Koroch, A.; Rodríguez, Y.; Sailer, J.; Teleshova, N. Algal and Cyanobacterial Lectins and Their Antimicrobial Properties. Mar. Drugs 2021, 19, 687. https://doi.org/10.3390/md19120687
AMA Style
Fernández Romero JA, Paglini MG, Priano C, Koroch A, Rodríguez Y, Sailer J, Teleshova N. Algal and Cyanobacterial Lectins and Their Antimicrobial Properties. Marine Drugs. 2021; 19(12):687. https://doi.org/10.3390/md19120687
Chicago/Turabian StyleFernández Romero, José Abel, María Gabriela Paglini, Christine Priano, Adolfina Koroch, Yoel Rodríguez, James Sailer, and Natalia Teleshova. 2021. "Algal and Cyanobacterial Lectins and Their Antimicrobial Properties" Marine Drugs 19, no. 12: 687. https://doi.org/10.3390/md19120687
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.